私たちは必須元素の中でも硫黄(S)に焦点をあてています。
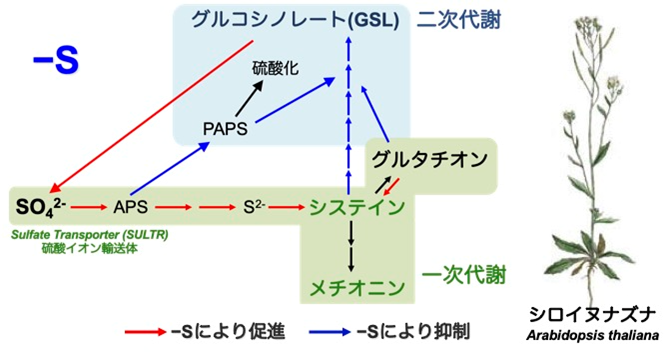
植物は、人間にとっても必要なアミノ酸であるシステインやメチオニンを、環境中の硫酸イオンから作り出します。無機の硫黄は酸性化をもたらすので、植物は自然界の硫黄サイクルにも大きな役割を果たしています。
植物が作り出す含硫化合物には、アミノ酸やタンパク質の他、酸化還元物質、補酵素や医薬として有用なものが多くあります。近年では、アブラナ科植物に多く含まれるメチオニン由来グルコシノレートの発ガン抑制効果が注目されています。
植物体内で硫黄の同化・代謝がどのように調節されているのかを理解できれば、作物生産性の向上、食を通した人間の健康増進、環境問題の解決に向けた手がかりになると考えています。
硫黄の同化と代謝の仕組み
硫黄の同化は、硫酸イオンの吸収から始まります。硫酸イオンの吸収に働くのが硫酸イオン輸送体(Sulfate Transporter, SULTR)です。吸収された硫酸イオンは、いくつかの酵素の働きにより還元され、システインへと同化されます(総説11)。その後、必須アミノ酸であるメチオニンや酸化還元反応に働くグルタチオンなど、硫黄を含む代謝物が生合成されます。
環境中の硫黄が不足すると、植物の体内では、硫黄同化に働く酵素の遺伝子発現が上昇します。逆に、グルコシノレートの生合成は減少します(論文11, 総説9, 12)。そうすることで、生命維持に必要なタンパク質の生産を維持するのです。
私たちは、この硫黄栄養の変化から遺伝子発現の変化に至る情報伝達系を研究することで、硫黄同化・代謝の仕組みとその調節機構を明らかにしようとしています。
硫黄不足に対する応答機構
植物はどのように硫黄不足を感知して代謝を調節するのでしょうか?
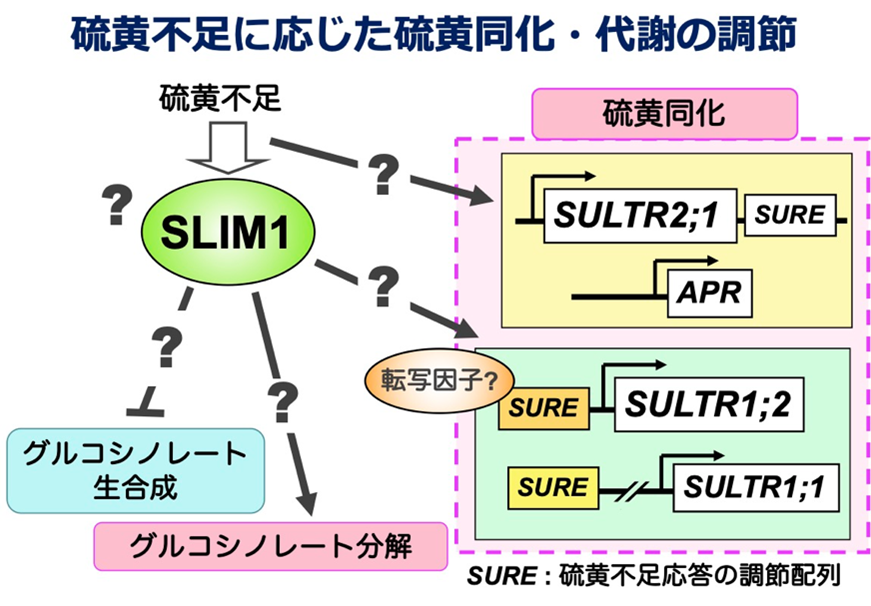
私たちは、硫黄不足に応じた硫黄同化の促進やグルコシノレート生合成の抑制を司る転写制御因子SLIM1を発見しました(論文16)。SLIM1の活性化にはC末端のアミノ酸配列が必要です(論文44)。
植物が硫黄不足をどう感知するのか? SLIM1がどのように働き出すのか? など、まだまだ多くの疑問が残されています。 この疑問を解決することで、硫黄同化を活性化する術を見出そうと考えています。
硫酸イオン吸収の調節
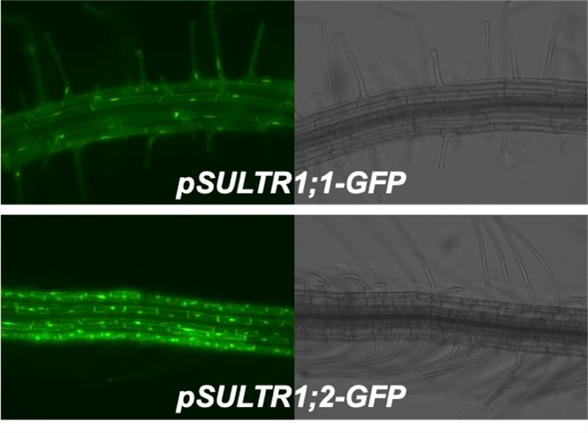
シロイヌナズナでは、硫酸イオン輸送体SULTR1;1, SULTR1;2 が硫酸イオンの吸収を担っています。
硫黄が不足すると、これらの遺伝子発現が促され、硫酸イオンの吸収活性が高まります。
これまでに、SULTR1;1の発現上昇を担う調節配列を決定しました(論文15)。また、この応答にタンパク質のリン酸化・脱リン酸化が関わることが分かりました(論文12)。SULTR1;2の調節配列の決定やこれらの発現を調節する転写因子を探索しています。
SULTR1;1, SULTR1;2は根の表面、表皮や皮層といった細胞層で働きます。表皮には、根毛が出る細胞と出ない細胞があり、SULTR1;1, SULTR1;2はそれぞれの細胞で発現します(論文32)。この綺麗な分かれ方に潜む意味にも興味をもっています。
メチオニン由来グルコシノレートの生合成・代謝の調節機構
アブラナ科植物に特有の含硫代謝物グルコシノレート(GSL)には、病害虫の忌避や、逆に特定の昆虫の誘因に働きます。メチオニンから作られるGSL (mGSL)には、発ガン抑制効果を示すものがあります。硫黄を貯蔵する役割もあります。
Sulfur Deficiency Induced (SDI)というタンパク質はmGSL生合成を抑制します。SDIの発現が硫黄不足により上昇し、mGSL生合成を促進する転写因子MYB28と相互作用することでその働きを抑え、mGSL生合成遺伝子の発現を抑えることでmGSL生合成を抑制します(論文26)。

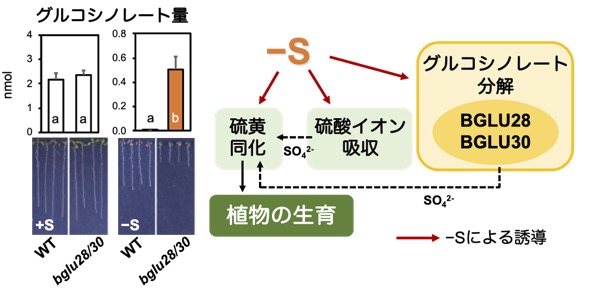
SDIやBGLU28, 30の発現が硫黄不足によって上昇する仕組みやSLIM1との関係を解析しています。
硫黄同化系を調節する新たな分子機構
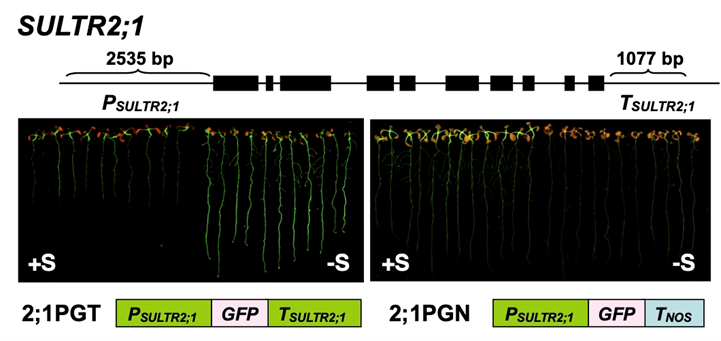
硫黄不足で発現の上昇する遺伝子の中には、SLIM1による制御を受けないものもあります。硫酸イオン輸送体SULTR2;1もその一つです。硫黄不足に応じたSULTR2;1の根における発現促進が3’領域に存在する配列によることを突き止めました(論文23)。この仕組みを明らかにできれば、SLIM1によらない新しい硫黄同化系の調節機構を理解できるのではないかと考えています。
他元素の分布や代謝に硫黄代謝が果たす役割
硫黄の代謝は、他の栄養素の代謝と協調して調節されます。生体内の窒素と硫黄の量は一定に保たれ、炭素や窒素栄養が不足していると硫黄不足への応答も起こりません(論文14)。
硫黄が不足すると、リン酸の吸収と地上部への輸送が促され、地上部のリン酸イオン量が増えます(論文38)。逆にリンの不足も硫酸イオンの輸送を変化させます。硫黄と鉄の間にも同様の関係があります。 このような栄養素間の相互調節の仕組みや植物にとっての意味にも興味を持っています。
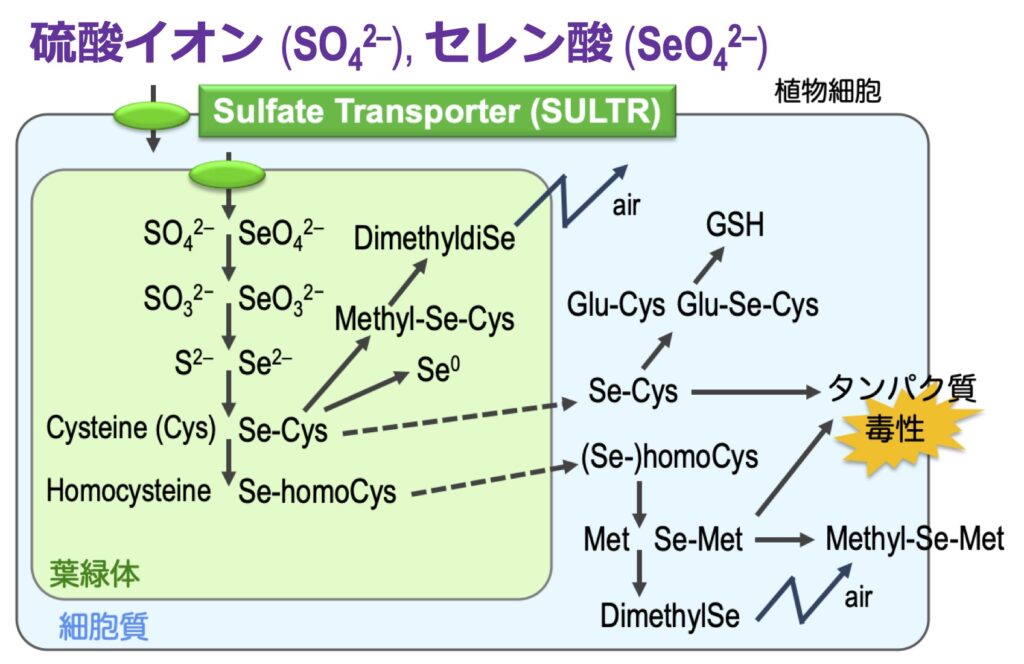
セレン(Se)は硫黄(S)と同族の元素で、植物体内での代謝機構も似ています。セレンは動物や微生物の必須元素であると同時に、高濃度では毒性を示します。セレンの高等植物での役割や、蓄積・毒性発揮の仕組みを硫黄同化・代謝の仕組みを切り口として明らかにしていこうと考えています。
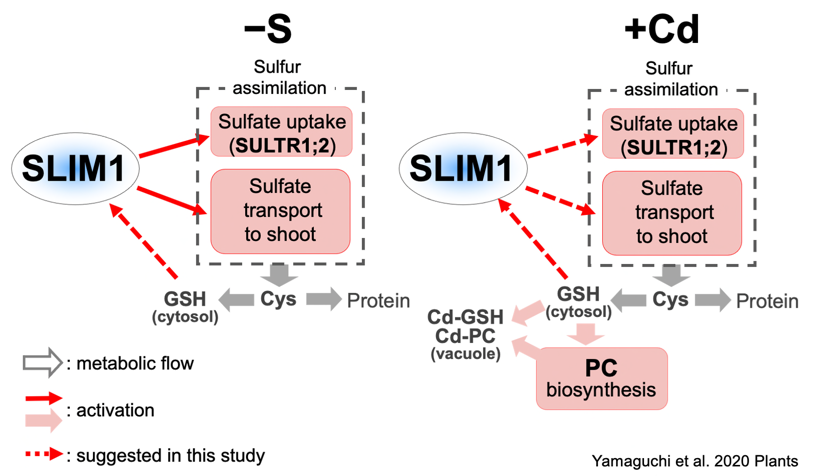
各種ストレスと硫黄
硫黄は植物のストレス耐性にも深く関わります。重金属の解毒に働くグルタチオンやファイトケラチンも硫黄を含みます。重金属にさらされた植物では、硫黄同化やファイトケラチン生合成が高まります(論文27)。このような状況では、硫黄が不足してもファイトケラチンの合成が優先され、SULTR1;2やSLIM1も寄与します(論文27, 29, 35)。
有用含硫化合物を高蓄積する・硫黄同化効率の高い植物の作出
大豆は植物由来のタンパク質源として重要ですが、栄養的にはシステインやメチオニンなどの硫黄を含むアミノ酸が制限となります。このため、植物を家畜の餌として用いる時にはメチオニンが加えられます。これまでに明らかにした事を活かして植物可食部中の硫黄量を増やす試みをしています。
硫黄同化・代謝を調節する化合物や環境条件
硫酸イオンの吸収や硫黄同化効率を変化させる環境要因や化合物の発見は、硫黄同化効率を高く保つ栽培方法や処理条件を見つけることに繋がります。
植物ホルモンであるサイトカイニンが、硫酸イオンの吸収を抑制することを見出しました(論文13)。また、ポルフィリン環の前駆物質であるγ-アミノレブリン酸が硫酸イオンの吸収やシステイン・グルタチオン量を増加させること、低濃度の塩が植物の生育と硫黄の蓄積を促すことを見つけました(論文41, 22)。
他にも硫黄の同化や代謝を調節する化合物や環境要因を見出すことで、情報伝達系を解明する手がかりを得ると同時に、生産現場に役立つ知見を得ようとしています。